研究プロセス
42 査読済み研究からの図表
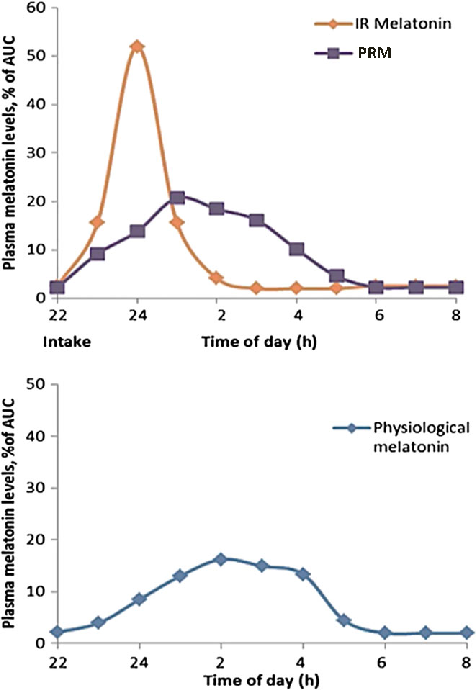
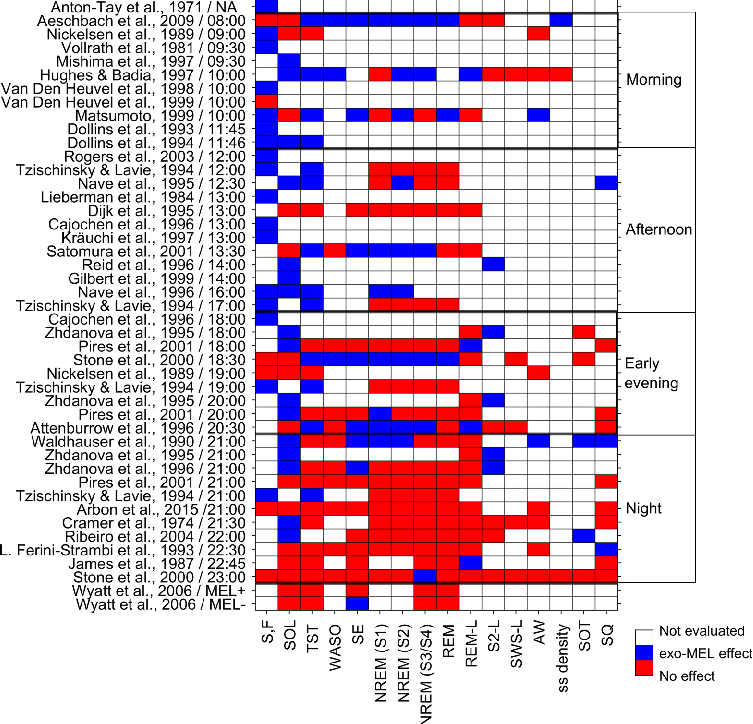
Exogenous melatonin administration influences circadian phase positioning, with the direction and magnitude of phase shifts depending on the timing of administration. Evening doses advance the circadian clock, while morning doses may cause phase delays.
New perspectives on the role of melatonin in human sleep, circadian rhythms …
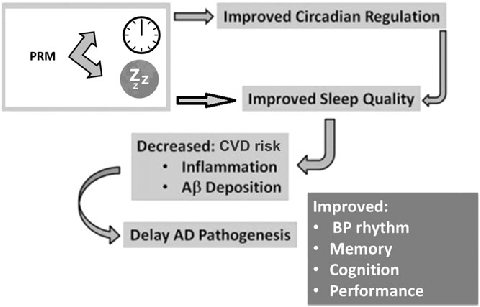
Clinical applications of melatonin extend beyond sleep induction to include circadian rhythm resynchronization in jet lag, shift work disorder, and delayed sleep-wake phase disorder. Dose-response relationships vary across these conditions.
New perspectives on the role of melatonin in human sleep, circadian rhythms …
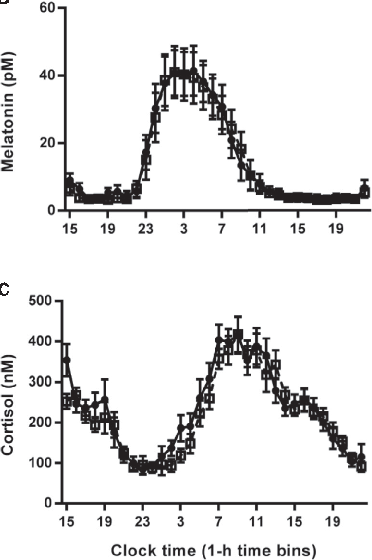
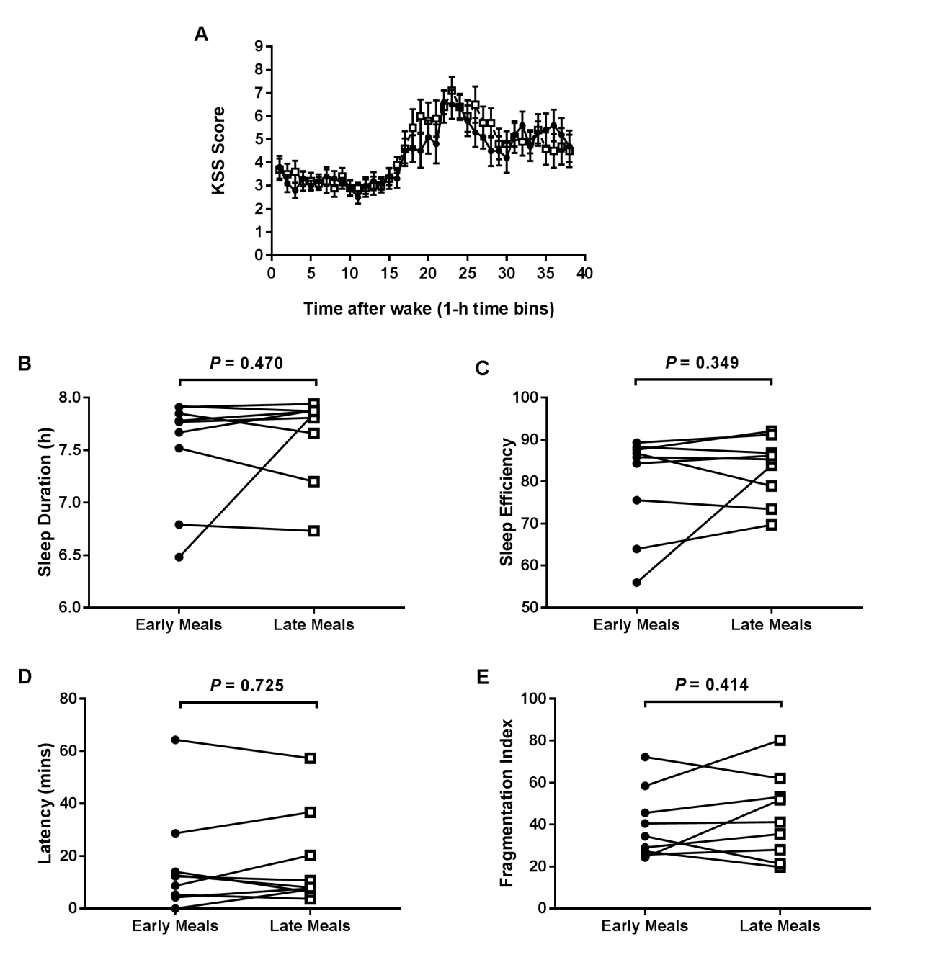
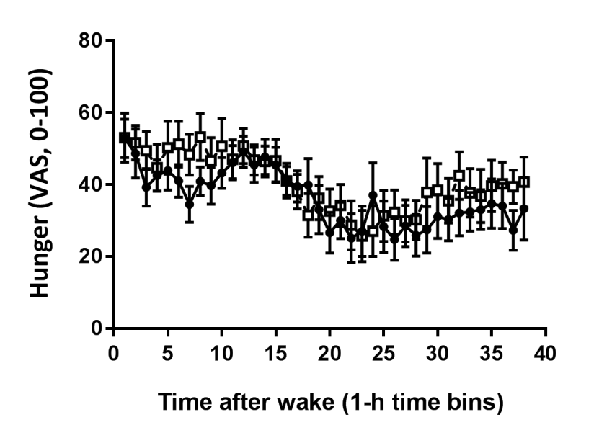
Hormonal response profiles to standardized meals are compared between normal and delayed meal timing conditions, revealing shifts in metabolic hormone rhythms.
Meal Timing Regulates the Human Circadian System.
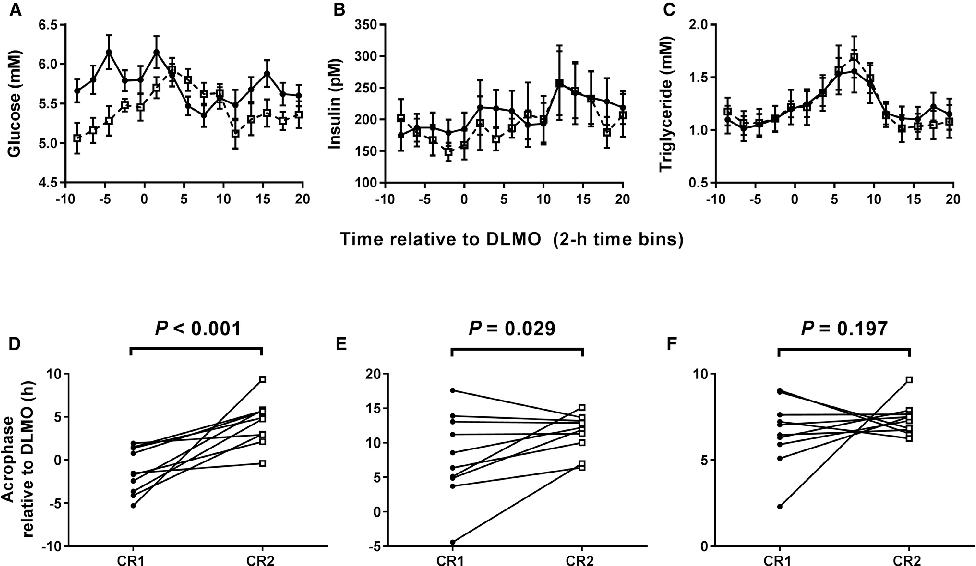
Average plasma glucose concentration under constant routine conditions is reduced following a 5-hour delay in meal times, suggesting meal timing significantly influences glucose metabolism rhythms.
Meal Timing Regulates the Human Circadian System.
Core body temperature rhythms are displayed across experimental conditions, demonstrating that meal timing shifts selectively affect peripheral but not central circadian markers.
Meal Timing Regulates the Human Circadian System.
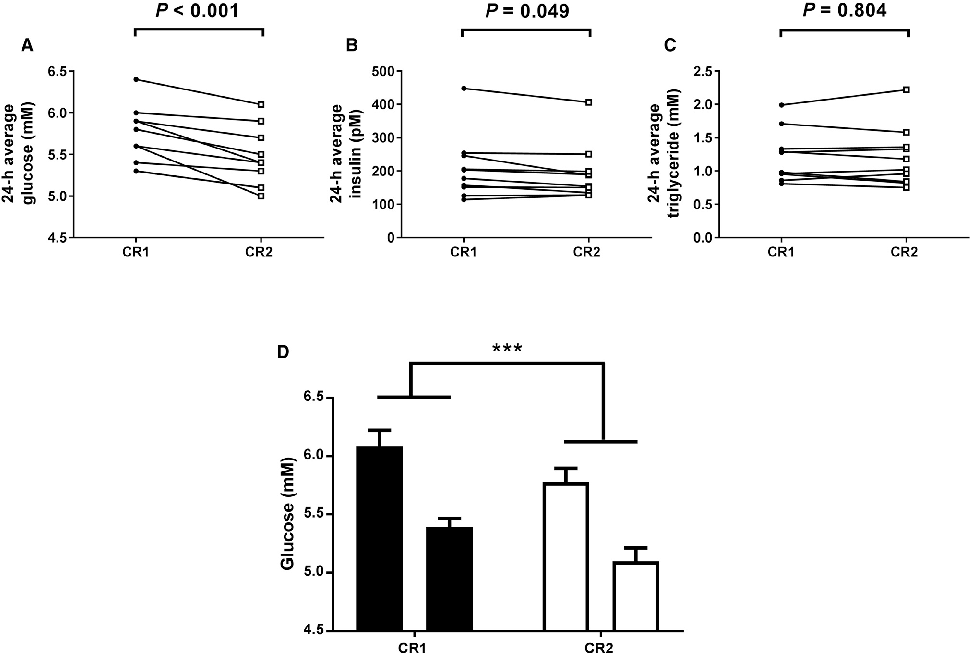
Plasma glucose concentrations during constant routine conditions show a significant reduction after participants experienced delayed meal times, indicating that meal scheduling influences circadian glucose regulation.
Meal Timing Regulates the Human Circadian System.
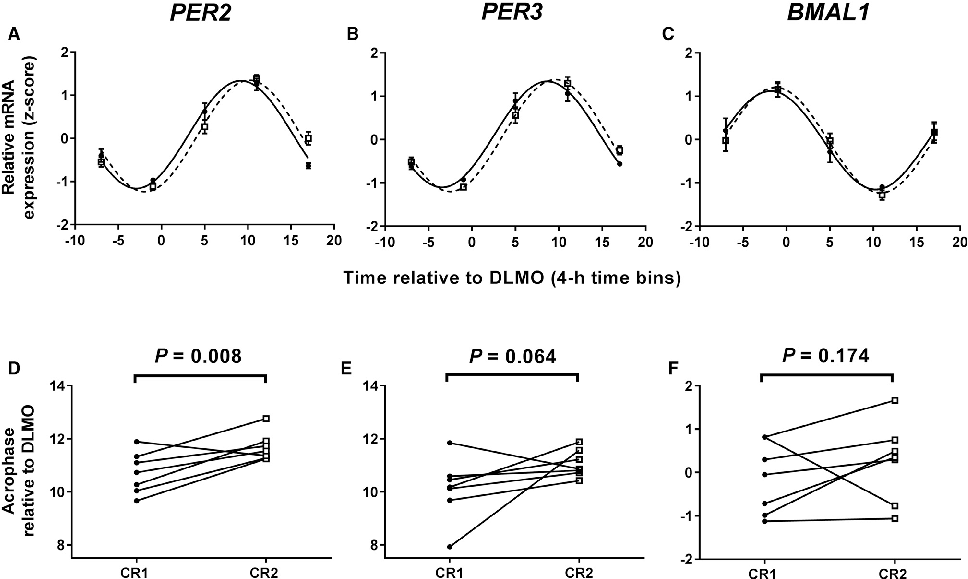
Clock gene expression patterns in adipose tissue are compared between the two meal timing conditions, revealing tissue-specific circadian phase shifts.
Meal Timing Regulates the Human Circadian System.
Per2 and other clock gene transcript levels measured from peripheral blood samples demonstrate delayed phase alignment consistent with the shifted meal schedule.
Meal Timing Regulates the Human Circadian System.
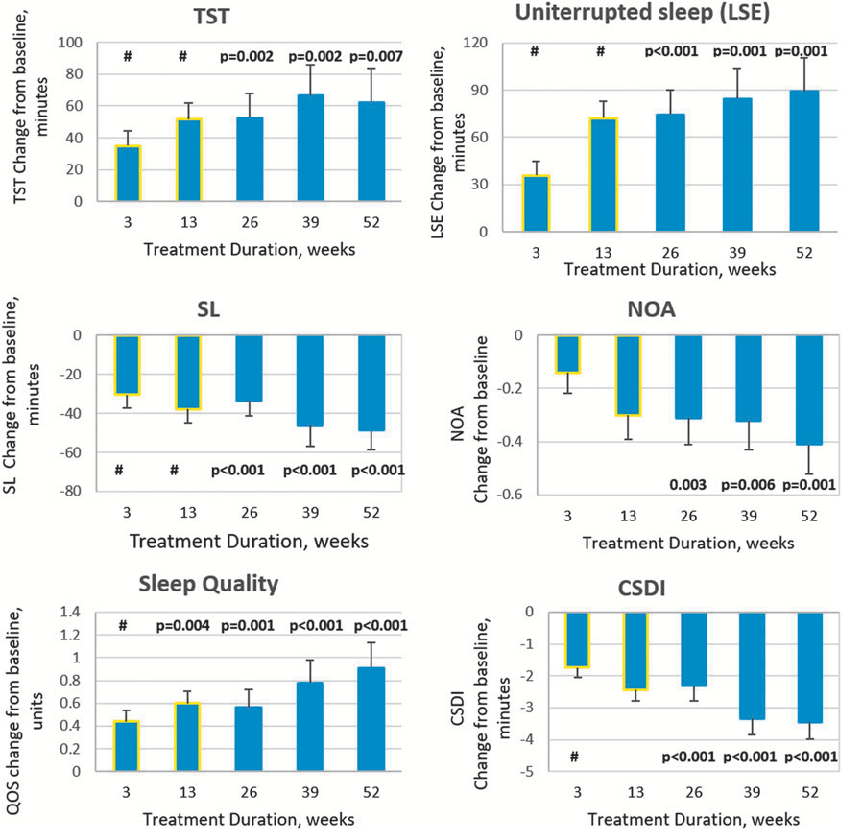
Efficacy outcomes showing changes in sleep parameters over the extended treatment period with pediatric prolonged-release melatonin in children with ASD. Long-term data indicate sustained improvements in sleep onset latency and total sleep time.
Long-Term Efficacy and Safety of Pediatric Prolonged-Release Melatonin for Insomnia in Children …
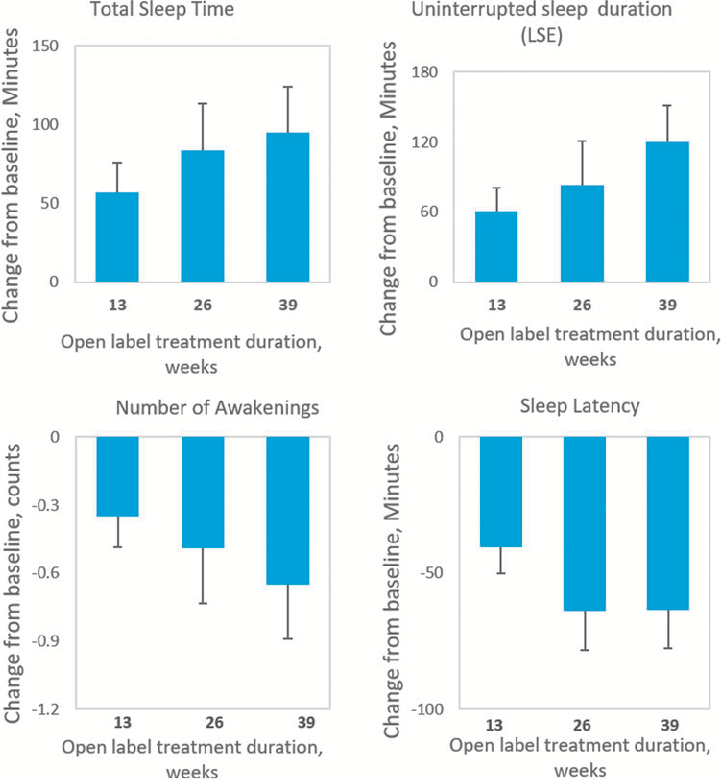
Safety assessment data from the open-label extension of PedPRM treatment in pediatric patients with autism spectrum disorder. Adverse event profiles suggest the prolonged-release melatonin formulation maintained an acceptable safety profile over the extended study period.
Long-Term Efficacy and Safety of Pediatric Prolonged-Release Melatonin for Insomnia in Children …
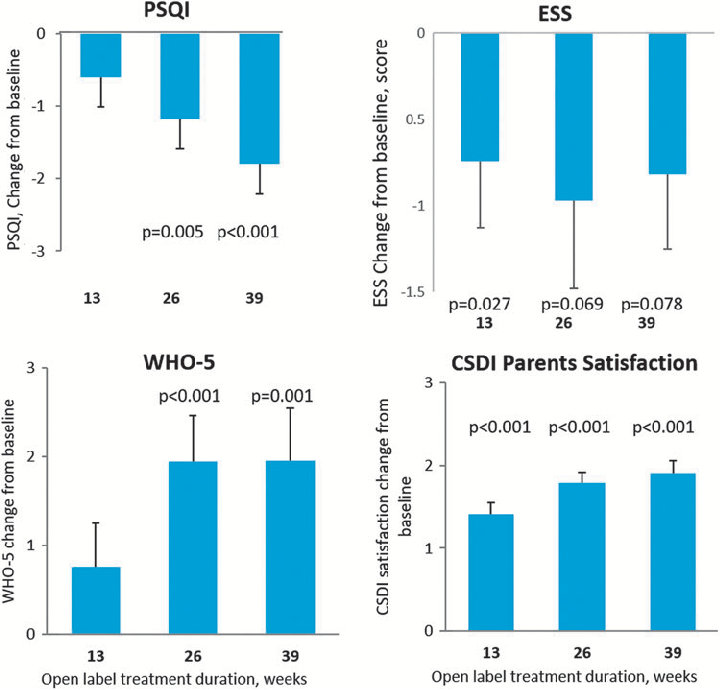
Summary of sleep quality measures or caregiver-reported outcomes during long-term PedPRM administration in children with ASD and comorbid insomnia. Results suggest continued therapeutic benefit with nightly melatonin use.
Long-Term Efficacy and Safety of Pediatric Prolonged-Release Melatonin for Insomnia in Children …
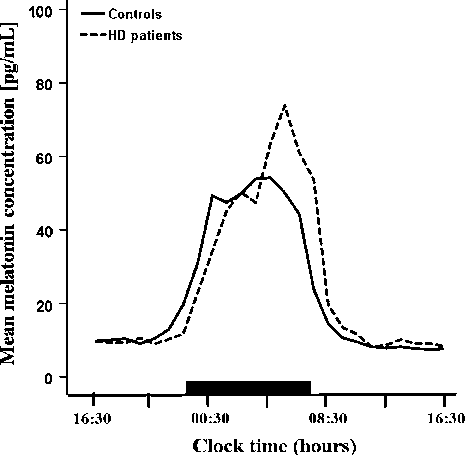
Mean 24-hour melatonin profiles in Huntington's disease patients and matched healthy controls are plotted, revealing a statistically significant delay of approximately 1.5 hours in the diurnal melatonin rise among HD patients (p = 0.048).
Delayed onset of the diurnal melatonin rise in patients with Huntington's disease.
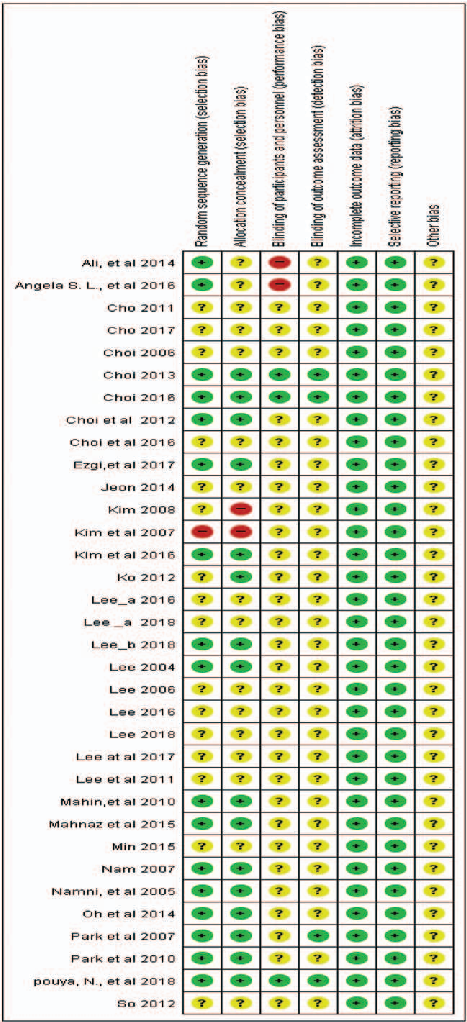
Risk of bias assessment across included studies evaluates methodological quality domains such as randomization, blinding, and outcome reporting in aroma therapy trials.
A systematic literature review and meta-analysis of the clinical effects of aroma …
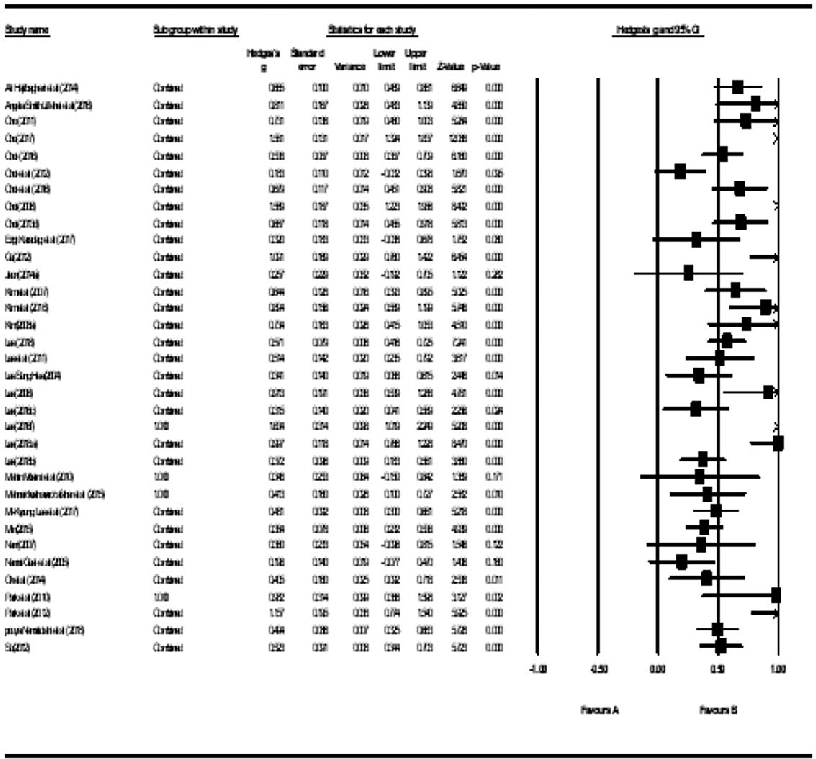
Traffic-light visualization of risk of bias for individual studies included in the aroma inhalation therapy meta-analysis, generated using RevMan software, reveals varying methodological quality.
A systematic literature review and meta-analysis of the clinical effects of aroma …
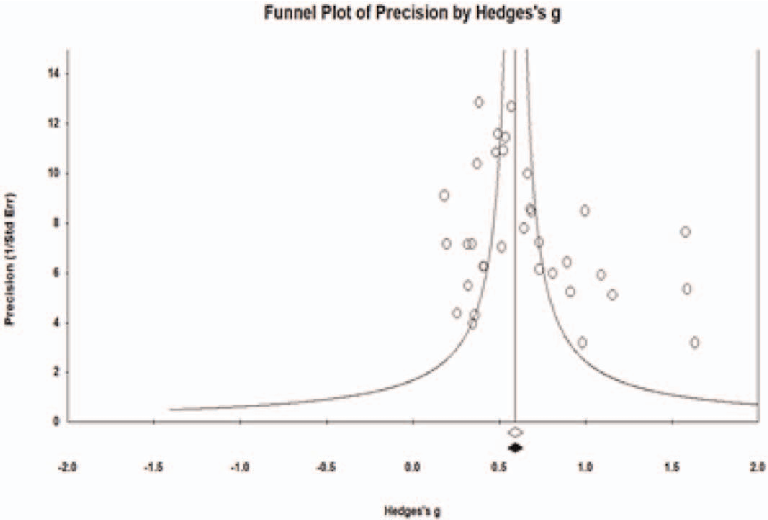
Funnel plot analysis assesses potential publication bias among studies included in the meta-analysis of aroma inhalation therapy for sleep problems, with symmetry indicating low risk.
A systematic literature review and meta-analysis of the clinical effects of aroma …
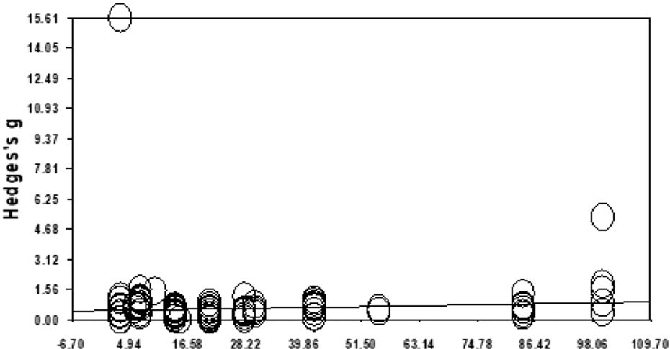
Meta-regression analysis examines whether the effectiveness of aroma inhalation therapy for sleep problems varies by program duration or publication year, revealing trends in the slope of the primary outcome.
A systematic literature review and meta-analysis of the clinical effects of aroma …
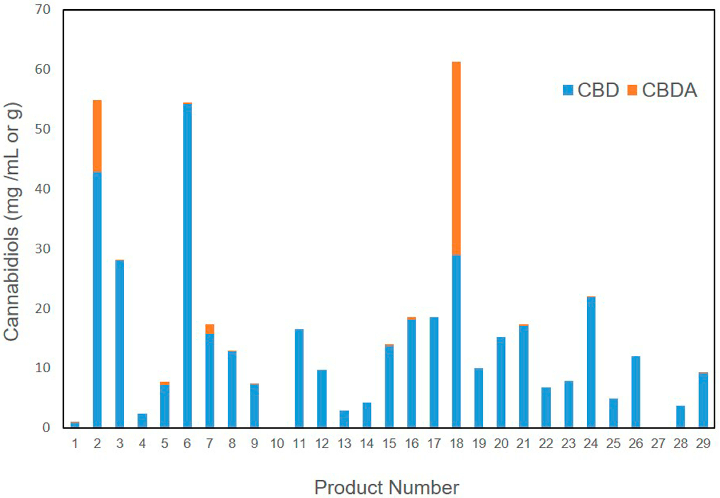
Overall quality assessment of commercial veterinary hemp supplements integrates cannabinoid accuracy, heavy metal safety, and terpene characterization. Substantial product-to-product variability underscores the need for standardized manufacturing.
Cannabinoid, Terpene, and Heavy Metal Analysis of 29 Over-the-Counter Commercial Veterinary Hemp …
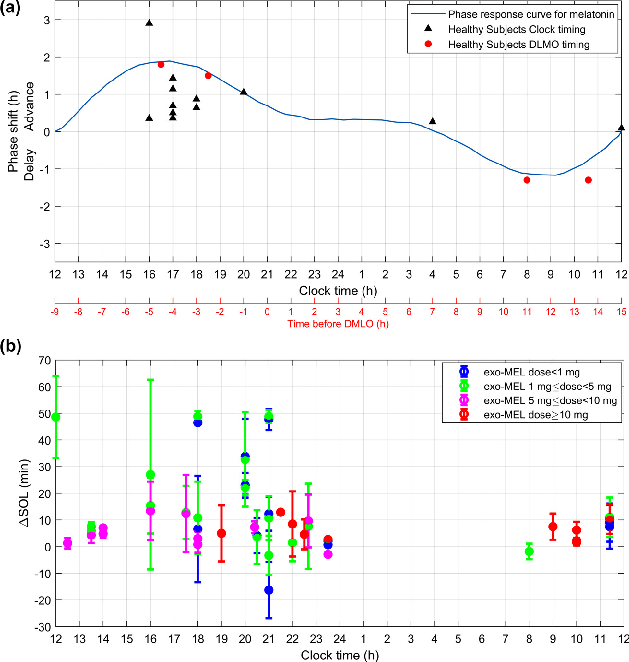
Dose-response analysis of melatonin supplementation on sleep parameters, indicating that lower physiological doses may be as effective as higher pharmacological doses for circadian rhythm regulation.
Melatonin as a Chronobiotic with Sleep-promoting Properties.
Circadian phase-response curve for exogenous melatonin, demonstrating how timing of administration determines whether it advances or delays the circadian clock.
Melatonin as a Chronobiotic with Sleep-promoting Properties.
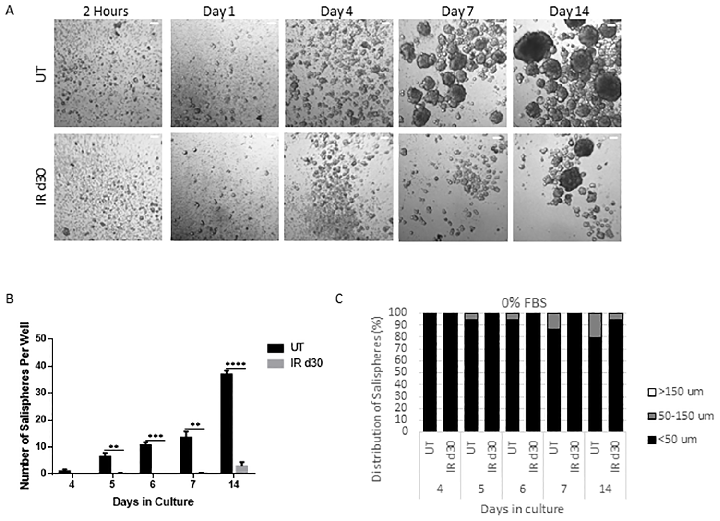
Sphere-forming efficiency of parotid-derived cells decreases significantly following a single 5 Gy radiation dose, quantified at day 30 post-irradiation.
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.
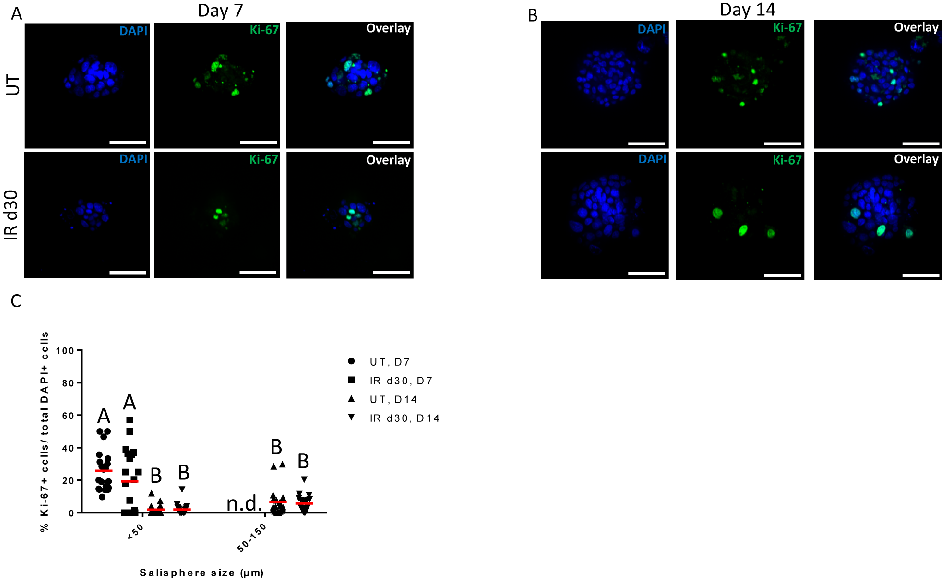
Proliferation rates of salisphere cultures from untreated and irradiated parotid glands are compared under serum-free conditions, showing similar growth kinetics despite radiation damage.
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.
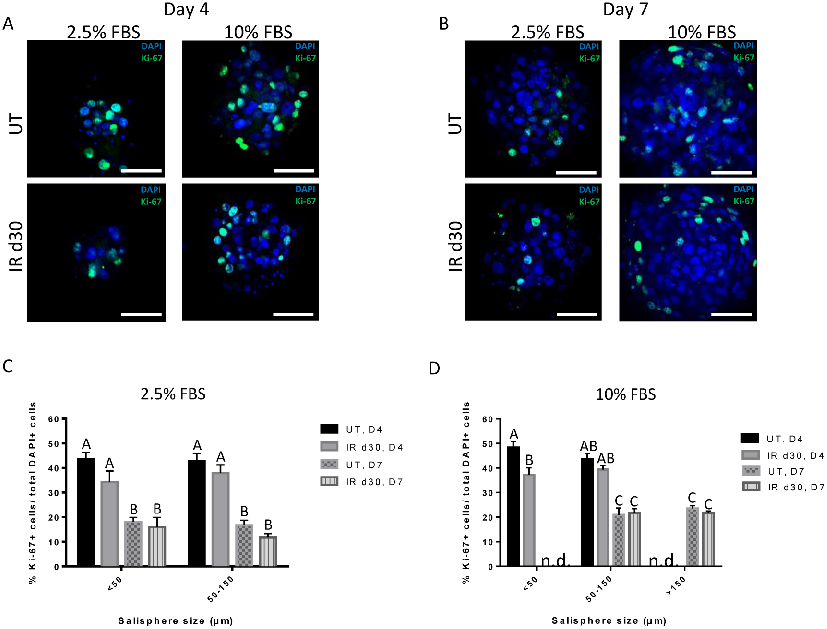
Proliferation assays confirm comparable growth rates between untreated and irradiated salisphere cultures supplemented with FBS, indicating that reduced sphere formation is not due to proliferation deficits.
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.
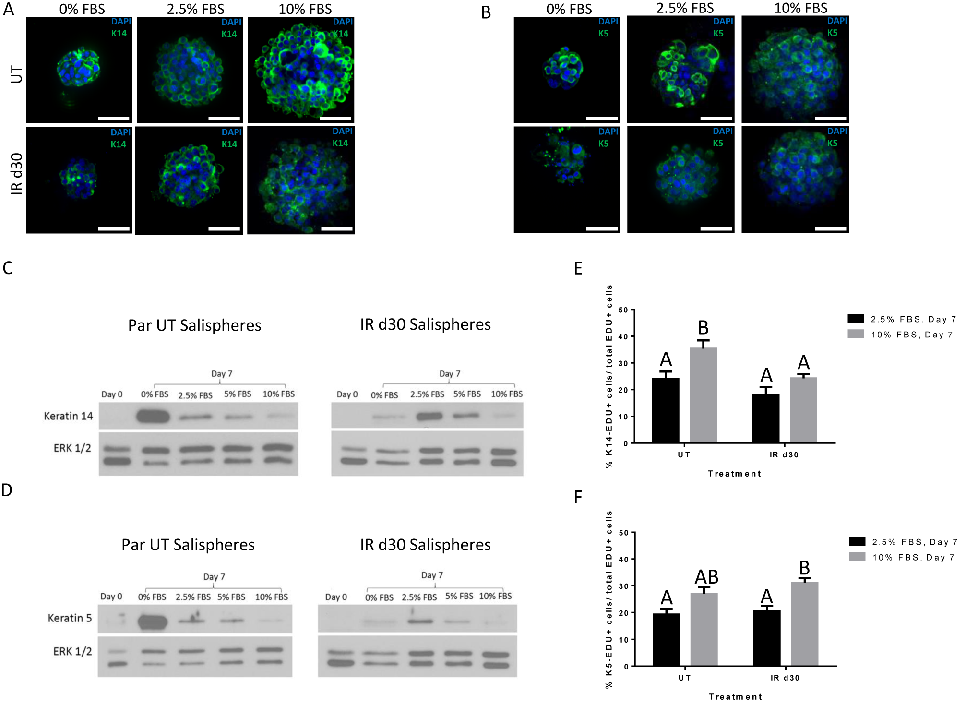
Expression of salivary stem and progenitor cell markers in salisphere cultures demonstrates that both untreated and irradiated cells maintain their progenitor phenotype.
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.
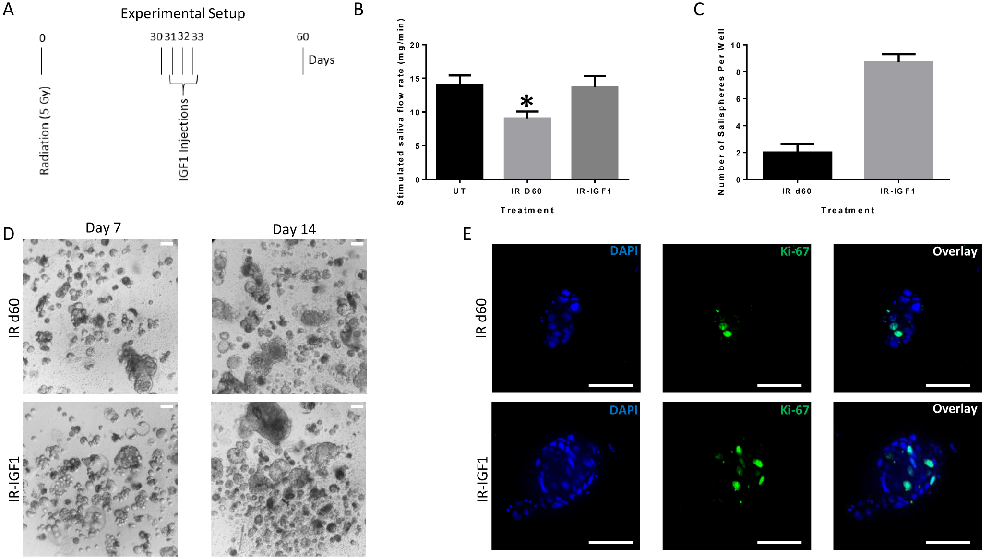
Post-radiation IGF-1 treatment significantly enhances sphere-forming efficiency of irradiated parotid cells, suggesting growth factor administration as a strategy to restore salivary function.
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.
2ページ中1ページ目